Click on images
to enlarge


Photographer: B.R. Maslin

Photographer: M.W. McDonald

Photographer: B.R. Maslin

Photographer: B.R. Maslin

Photographer: M.W. McDonald
, LSWE 7061, lab photo by Fiona McCallum ADJUSTED.jpg)
Seed from one herbarium voucher. Scale in mm. Photographer: F. McCallum.
Botanical name
Acacia colei var. ileocarpa M.W. McDonald & Maslin, Nuytsia 11: 219, fig. 1 (1997)
Common name
Curly-podded Cole's Wattle
Description
Shrubs or small trees 1.5-5 m tall, single- or multi-stemmed, stems often about 5-10 cm in diameter at breast height (to 50 cm on largest plants), crowns open and somewhat spindly under natural conditions but rather dense and bushy in disturbed, open sites such as along road verges, the main branches ascending to erect. Bark grey, smooth except finely longitudinally fissured at base of mature stems. Branchlets acutely angular and often orange at extremities, soon terete, densely sericeous with minute, closely appressed- straight, silvery hairs. New shoots pale yellow sericeous and often tinged brown by resin hairs when first initiated, hairs soon aging silvery. Phyllodes normally obliquely narrowly elliptic, narrowed at both ends, 7-19 (-21) cm long, (1-) 2-3 cm wide, to 5 cm wide on young plants, ascending to erect but some wide-spreading with age, ±straight but normally shallowly recurved at apices, rarely shallowly falcately recurved over their entire length, sericeous hairs as on branchlets (indumentum especially evident on young phyllodes and becoming sparser with age), silvery green, silvery grey-green or silvery bluish green; with 3 (-4) main longitudinal nerves (nerves becoming obscure towards phyllode apex and some confluent and ±contiguous with the lower margin at base), the minor nerves numerous and rather close together, anastomosing to form a longitudinally-orientated reticulum; the apex terminated by a knob-like callose point 0.5-1 mm. Gland absent from apex of phyllode. Inflorescences rudimentary 2-headed racemes with axes to 0.5 mm long; peduncles (2-) 3-5 (-10) mm long, appressed-hairy, the indumentum sometimes sparse; spikes large and showy, 3-6 cm long, bright golden, the flowers close together in the bud but becoming sub-densely arranged at anthesis. Flowers 5-merous; calyx very shortly dissected; petals hairy. Pods raised over the seeds and moderately constricted between them, 2.5-4 mm wide, thinly coriaceous-crustaceous, tightly and irregularly coiled and twisted, dehisced valves somewhat entangled and often persisting on the plants as conspicuous, ±spherical clumps (resembling birds' nests from a distance), reddish brown. Seeds longitudinal in the pods, obloid or sometimes obloid-ellipsoid or obloid-ovoid, 3-3.5 (-4) mm long, glossy, black; aril bright yellow.
Characteristic features
Shrubs or small trees. Branchlets acutely angular and often orange at extremities, densely sericeous. New shoots pale yellow sericeous aging silvery sericeous. Phyllodes large (mostly 7-19 x 2-3 cm), mostly ascending to erect, ±straight but normally shallowly recurved at apices, with a distinct silvery sheen (overall colour is silvery green, silvery grey-green or silvery bluish green), with 3 (-4) main longitudinal nerves (some confluent and ±contiguous with the lower margin at base of phyllode), minor nerves numerous and anastomosing to form a longitudinally-orientated reticulum; the apex terminated by a knob-like; no gland at apex of phyllode. Spikes large (3-6 cm long) on short peduncles (mostly 3-5 mm); petals hairy. Pods tightly and irregularly coiled and twisted. Seeds black, the aril bright yellow.
Distribution and ecology
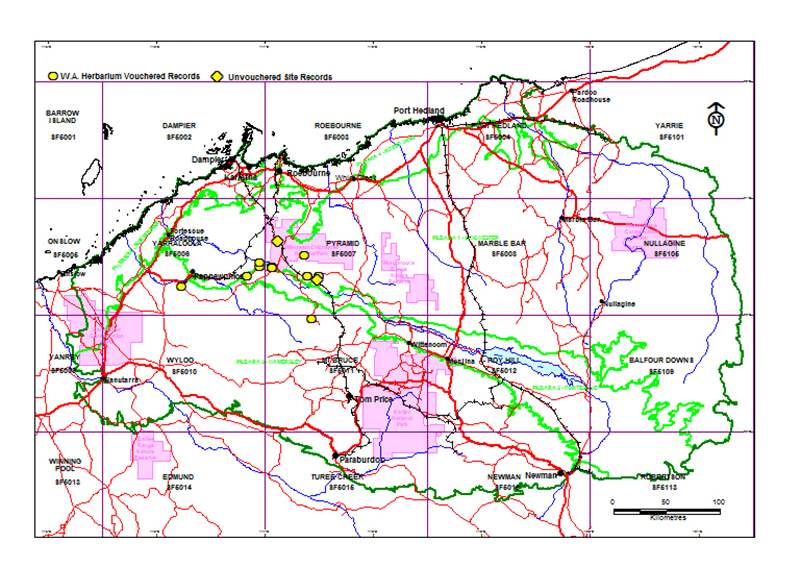
Scattered in the southern Kimberley region of Western Australia where it extends from near Derby eastward to the western extremity of Northern Territory at the Negri River; there is also a disjunct occurrence in the Pilbara where it has a restricted to the Panawonica - Millstream area. Although var. colei and var. ileocarpa occasionally co-occur in the Kimberley this seems not to be the case in the Pilbara although they grow do within a few kilometers of one another in some areas. Variety colei is far more common in the Pilbara than is var. ileocarpa. In the Pilbara var. ileocarpa favours stony spinifex plains in slightly alkaline sandy loam (pH 7.5-8.5) associated with water course and diffuse drainage lines where it forms rather dense but localized populations.
Flowering and fruiting period
Variety ileocarpa flowers mainly in June and July and pods with mature seeds occur between September and November.
Variation
Plants of var. ileocarpa display a range of variation in phyllode size and shape similar to, or perhaps even greater than, var. colei. Variety ileocarpa phyllodes are commonly 10-25 mm wide and as such tend to be slightly narrower (and often more attenuate) than those of var. colei. However, plants with some wider than normal phyllodes (to 40 mm across and clearly within the range of variation for var. colei) occur scattered throughout the geographic range of the variety. Further field work is required to ascertain if these specimens represent collections from young plants because many taxa in the group to which A. colei belongs tend to produce larger than normal phyllodes during their early growth phase.
Taxonomy
Acacia colei comprises two varieties, var. colei and var. ileocarpa, both of which occur in the Pilbara.
Affinities
In the absence of pods it is normally difficult to distinguish between var. colei and var. ileocarpa. The undehisced pods of var. ileocarpa are irregularly coiled or twisted (very similar to those found on related species A. holosericea , which is rare in the Pilbara, and A. neurocarpa, which does not occur in the Pilbara) whereas in var. colei the undehisced pods are curved (sometimes into an open circle). Care needs to be exercised when using this pod curvature character because following dehiscence the pods of both varieties become somewhat twisted and entangled and often persisting on the plants as conspicuous, ±spherical clumps. It is therefore important to inspect pods prior to their dehiscence when using this character to identify the varieties. Another, but less reliable, way to separate the varieties is by the colour of their terminal branchlets which is often orange in var. ileocarpa and brown in var. colei. There is also a tendency for var. ileocarpa to have slightly narrower phyllodes than those of var. colei, however, there is a large overlap in phyllode width between the varieties. As already noted, var. colei is far more common in the Pilbara than is var. ileocarpa. Acacia holosericea is distinguished from var. ileocarpa by its generally straighter phyllodes with a different nervation pattern and some of which have a small gland at their apices.
Notes
Both varieties of A. colei have shown potential for multipurpose use in tropical dry-zones of West Africa. Recently their seeds have been used as an alternative source of human food in these regions and the seeds are ground-up to produce a flour which can be incorporated into local recipes (Rinaudo et al. 1995). Today farmers near Maradi, Niger, have shown a clear preference for var. ileocarpa over var. colei as it has better harvesting characteristics (Rinaudo and Thomson 2006). In var. ileocarpa the seeds are held firmly in their coiled pods, hence ripe pods can be left on the tree longer without seed loss. Pickers can wait for a high proportion of the pods to ripen before harvesting, spending less time on repeat harvests. Additionally, even though the pods hold seed longer, once fully dry, a light beating with a stick will separate seed from pods. In var. colei the pods ripen unevenly, and seed begins to drop from pods within a day or so of ripening. Thus, pickers must return daily to harvest newly ripened pods.
The cytogenetics of progeny from plants grown in Maradi, Niger, show that var. ileocarpa is hexaploid (2n = 78) (Helen Stace, pers. comm.). This is the same ploidy level as var. colei reported by Moran et al. (1992).
Conservation status
Not considered rare or endangered.
Origin of name
The varietal epithet is derived from the Latin - ileum (last part of the small intestine, alluding to the twisted and coiled state of its pods) and the Greek - carpa (fruit).
References
Moran, G.F., Thomson, L., Grant, J. and Bell, C. (1992). Distribution of genetic variation within two dry-zone Acacia species and implications for their genetic improvement. pp. 74-81. In: A.P.N. House and C. Harwood (eds) Australian Dry-zone Acacias for Human Food. Proceedings of a workshop held at Glen Helen, Northern Territory, Australia, 7-10 August, 1991. pp. 151. (Australian Tree Seed Centre, CSIRO Division of Forestry: Canberra.)
Rinaudo, T., Burt, M. and Harwood, C. (1995). Growth and seed production of Australian Acacia species at Maradi, Niger. ACIAR Forestry Newsletter, No. 19. (Australian Centre for International Agricultural Research: Canberra.)
Rinaudo, T. and Thomson, L. (2006). Multipurpose useage of Australian Acacias in sub- Saharan Africa. WorldWideWattle website. http://www.worldwidewattle.com/infogallery/utilisation/sehel.php [Accessed 2009]