Acacia neurocarpa A.Cunn. ex Hook., Icon. Pl . 2: t. 168 (1837)
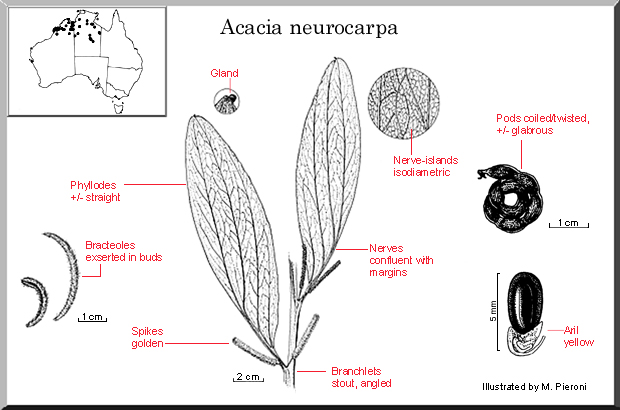
Shrub or tree 3–8 m high. Branchlets stout and prominently angled for a considerable distance below apex, velutinous to sericeous. Young shoots silvery sericeous. Phyllodes ascending to erect, obliquely narrowly elliptic, mostly 12–25 cm long and 3–5.5 cm wide, with unequal base, the mucro 0.5–1.5 mm long, dull green to greyish green, sericeous or velutinous, the normally 3 prominent veins per face contiguous and confluent with adaxial margin at base, prominently and closely reticulate; glands basal and at base of mucro on at least some phyllodes. Inflorescences simple; peduncles 4–10 mm long, puberulous; spikes 4–7 cm long, golden; bracteoles linear, 2–4 mm long, evident and often spreading in buds. Flowers mostly 5-merous; sepals 2/3–3/4-united; petals puberulous. Pods rather tightly and often irregularly coiled, 3–4 mm wide, thinly coriaceous-crustaceous, subglabrous, persisting as entangled clumps following dehiscence. Seeds longitudinal, oblong, 3–5 mm long, glossy, dark brown to black; aril bright yellow.
Common from the Dampier Penin., N of Broome, W.A., eastwards through the N.T. where the distribution is more scattered, to Springvale near the N.T./Qld border. Grows in sand along seasonally wet watercourses, in woodland or riparian forest, often in association with Melaleuca sp.
Until recently A. neurocarpa was confused with A. holosericea , fide B.R.Maslin & L.A.J.Thomson, Austral. Syst. Bot . 5: 729–743 (1992). Putative hybrids occur between these species in the Bungle Bungle Natl Park, W.A. (e.g. K.A.Menkhorst 524 , DNA, PERTH) and in the Nicholson R. area, N.T. (e.g. T.Henshall 353 , DNA). Although more work is needed in order to clarify the differences between the two species, A. holosericea is generally recognised by its its shorter, spathulate bracteoles, normally narrower phyllodes with a more open reticulum comprising vein-islands that tend to be more elongated and aligned subparallel to the main veins, and more slender branchlets (3–6 mm wide and terete or slightly angled at 20–30 cm below the apex, compared with 5–12 mm wide and acutely angled in A. neurocarpa ).
The hexaploid species A. colei appears to have evolved as an allopolyploid between A. neurocarpa (diploid) and A. cowleana (tetraploid), fide G.F.Moran et al . (1992), in A.House & C.Harwood (eds), Austral. Dry-Zone Acacias Human Food (CSIRO, Canberra).
As discussed by B.R.Maslin & L.A.J.Thomson, loc. cit ., it is almost certain that Allan Cunningham applied the name A. neurocarpa to the species known today as A. dunnii . However, when formally describing A. neurocarpa W.J.Hooker confounded two species under this name. In the interest of nomenclatural stability B.R.Maslin & L.A.J.Thomson, loc. cit ., lectotypified A. neurocarpa to exclude A. dunnii , even though the epithet is inappropriate for the species now bearing this name.
Type of accepted name
‘Acacia neurocarpa. Camb. Gulf’ [this specimen is probably part of the A.Cunningham 478 collection said to have been collected from Port Keats, N.T. and Cambridge Gulf, W.A., Sept. 1819]; lecto: K, fide B.R.Maslin & L.A.J.Thomson, Austral. Syst. Bot . 5: 736 (1992); paralecto: see A. dunnii .
Synonymy
Acacia holosericea var. neurocarpa (A.Cunn. ex Hook.) Domin, Biblioth. Bot . 89: 270 (1926). Type: as for accepted name.
Illustrations
W.J.Hooker, Icon. Pl . 2: t. 168 (1837), as to fruiting specimen; B.R.Maslin & L.A.J.Thomson, Austral. Syst. Bot . 5: 734, fig. 2 (1992).
Representative collections
W.A.: Talbot Ck, 3 km SE of Mt Hart HS, D.J.Edinger 400 (K, NSW, PERTH); L. Argyle tourist village, Kununurra, K.F.Kenneally 19166 (CANB, K, L, PERTH); c. 7 km SW of Chattur Bay, Dampier Penin., B.R.Maslin 4935 (BRI, PERTH). N.T.: 30 km ESE of Hatches Ck Store, N.Forde 813 (DNA); Springvale, A.Nichols 629 (BRI).
(BRM)

The information presented here originally appeared on the WATTLE CD-ROM which was jointly published by the Australian Biological Resources Study, Canberra, and the Department of Parks and Wildlife, Perth; it was produced by CSIRO Publishing from where it is available for purchase. The WATTLE custodians are thanked for allowing us to post this information here.