Generic classification
Acacia is a member of tribe Acacieae within subfamily Mimosoideae of the family Leguminosae (the Pea Family). The Mimosoideae (which is sometimes treated as a distinct family, the Mimosaceae) contains about 50–60 genera that are distributed throughout tropical, subtropical and warm-temperate regions of the world (Elias 1981 and Cowan 1998). Tribe Acacieae contains just two genera, the large cosmopolitan genus Acacia (containing about 1380 species, rendering it the largest genus within the Mimosoideae) and the monotypic genus Faidherbia that occurs in Africa and the Middle East. (Note: Faidherbia is sometimes classified as a member of the closely related tribe Ingeae, see Polhill 1994; additional information on the classification and phylogeny of Faidherbia is given in Maslin & Stirton 1997 and Maslin et al. 2003). As summarized by Maslin et al. (2003) many uncertainties exist not only with the status of tribe Acacieae, especially in relation to tribe Ingeae and to a lesser extent tribe Mimoseae, but also with the definition, classification and phylogeny of Acacia itself. A cladistic analysis of the Mimosoideae based on characters of morphology, pollen, chemistry and anatomy is provided in Chappill & Maslin (1995) and cladistic analyses based on a synthesis of chloroplast DNA sequence data is provided by Luckow et al. (in press).
Recent studies of the phylogeny and generic status of Acacia using cladistic analyses of morphological and chloroplast DNA sequence data have been especially informative in developing a better understanding of the group. Of particular relevance are papers by Chappill & Maslin (1995), Grimes (1999), Robinson & Harris (2000), Miller & Bayer (2000, 2001 and 2003) and Luckow et al. (in press). These new data clearly show that the genus as presently defined (i.e. Acacia sens. lat.) is not monophyletic and most likely will be split into a number of separate genera at some future date (see Maslin et al. 2003 for summary). However, it is not entirely certain at present how many genera will be recognized, or indeed, what names will be applied to at least some of them.
The classification history of Acacia and nomenclatural implications associated with splitting the genus is given elsewhere on WorldWideWattle; a simplified version of these issues, and in particular, the nomenclatural impacts, is also given.
The following synopsis outlines the current higher-order classification of the Acacia.
The circumscription of Acacia has been relatively stable since about the mid-19th century, following a series of papers between 1842 and 1875 by the British botanist, George Bentham. The genus was first described by Philip Miller in 1754 and until 1842, when Bentham clearly defined it’s limits (by restricting the name Acacia to Mimosoid plants having numerous free stamens), a number of species which are now referable to genera within tribes Ingeae and Mimoseae, had been referred to it. In 1875 Bentham published his final (worldwide) classification of Acacia and this remained in place for about 100 years. The first major rearrangement of Bentham’s scheme was undertaken by the French botanist, Jacques Vassal in 1972. Vassal divided Acacia into three subgenera, Acacia, Aculeiferum and Heterophyllum (=Phyllodineae), and this work assisted in providing the conceptual framework for Pedley’s (1986) formal split of the genus into three genera. Initially Pedley (1978) viewed Acacia as comprising three large subgenera, as Vassal had done previously. However, in 1986 Pedley raised the rank of these groups to that of genus, Acacia, Senegalia and Racosperma respectively. Although it was generally accepted at the time that Acacia comprised a number of disparate groups, Pedley’s proposal was not widely adopted by the botanical community. Of primary concern was the widespread nomenclatural disruption that would ensue from splitting this enormous cosmopolitan genus. It was considered that more comprehensive information was needed in order to make informed decisions, and in particular, the need for broad-based comparative studies of Acacia that included genera from tribes Ingeae and Mimoseae was identified. Discussion of the reasons for not accepting Pedley's generic classification are outlined in Maslin (1987 and 1988, the latter paper was reproduced in 1989). Pedley published a defence of his scheme in 1987 and 1989.
Since 1986 there has been an accumulation of much new data derived from both morphological and molecular genetic studies, and this has led to a better understanding of the classification and phylogeny of Acacia. A summary of these new data is given in Maslin et al. (2003). This new work has confirmed that subg. Acacia and subg. Phyllodineae are monophyletic, however, subg. Aculeiferum is not monophyletic. Maslin et al. (2003) recognize three monophyletic assemblages within this last subgenus and suggest that at least five genera can be recoginzed within Acacia sens. lat. (species included in each of these genera are shown elsewhere on WorldWideWattle):
- AcaciaA (based on Acacia subgenus Acacia; a pantropical group containing about 163 species).
- Senegalia sens. str. (based on Acacia subgenus Aculeiferum; a pantropical group containing 203 species).
- Acaciella (based on Acacia subg. Aculeiferum section Filicinae; a group of 15 species confined to the Americas).
- Genus x (an undescribed genus based on a group of 13 species related to Acacia coulteri; confined to the Americas).
- RacospermaA (based on Acacia subgenus Phyllodineae; a group largely confined to Australia and containing 987 species).
AThe names Acacia and Racosperma as used here (and in Table 1) are those that would apply under the provisions of the International Code of Botanical Nomenclatrue by the application of currently designated types for these two genera. However, a proposal to retypify Acacia is currently in progress (Maslin & Orchard 2003) and if successful the following name changes would occur: Racosperma would be called Acacia and Acacia would be called Vachellia.
The classifications of Bentham (1875), Vassal (1972), Pedley (1978 and 1986) and Maslin et al. (2003) are shown in Table 1.
Note: Sec = Section, Ser = Series, Sser = Subseries, Sg = Subgenus
1Formalized in Guinet and Vassal (1978)
2The type of subseries Brunioideae is referable to sec Phyllodineae, however, most taxa that Bentham included in this group are referable to sec Lycopodiifoliae; none of these species was included in Vassal’s classification.
3Section Pulchelloidea included species from Bentham Subseries Pulchellae, Alatae, Continuae, Calamiformes, Plurinerves and Uninerves.
4An undescribed genus based on the species of the Acacia coulteri group (see Jawad et al. 2000).
Classification of Australian Acacias of subgenus Phyllodineae
Acacia is the largest genus of vascular plants in Australia and a majority of the 975 described, accepted species are contained in subgenus Phyllodineae. There are only 19 species of the subgenus (contained in sections Juliflorae and Plurinerves) that occur outside the continent and these are found in Pacific region (east to Hawaii), Asia (north to the Phillipines) and on Madagascar and the Mascarene Islands in the Indian Ocean off the east coast of Africa. Except for A. pubirhachis which occurs in New Guinea these extra-Australian species are discussed in Pedley (1975).
All recent cladistic studies based on both molecular and morphological data support subgenus Phyllodineae as monophyletic (Chappill & Maslin 1995, Miller & Bayer 2000 and 2001 and Robinson & Harris 2000). Although the phylogenetic relationships of the subordinate groups within the subgenus have not yet been satisfactorily resolved (see Maslin et al. 2003 for discussion) it is expected that on-going nuclear and chloroplast DNA studies will ultimately provide data to enable the construction of a meaningful classification of this group, one which is phylogetically based and which will accommodate the species in a hierarchial system containing an appropriate number of lower-order categories.
Currently the most generally used classification of subgenus Phyllodineae is that of Pedley (1978) in which seven sections are recognized, namely, section Alatae, Botrycephalae, Juliflorae, Lycopodiifoliae, Phyllodineae, Plurinerves and Pulchellae (Table 1). As discussed by Maslin (2001) this scheme represents a pragmatic attempt at rationalizing earlier classifications by Bentham (1842, 1864, 1875) and Vassal (1972), and offers a reasonably convenient way of arranging species into higher-order groups, but not all species are well-accommodated by these sections. Nevertheless, this scheme is a useful framework for the present discussions.
Recent cladistic, molecular and morphological studies suggest that, with the possible exception of the relatively small sections Lycopodiifoliae and Pulchellae, the sections of subgenus Phyllodineae (which contain about 95% of the species) are either paraphyletic or polyphyletic. Nevertheless, these studies and a range of other investigations conducted over the past few decades do reveal that some general phylogenetic trends are beginning to emerge (see Maslin et al. 2003 for discussion). The more significant of these are noted in the following paragraphs (further details are given in the sectional discussions below).
- The general trend in the recent morphological and DNA studies is for certain ‘uninerved’ phyllode-bearing species with elongated racemose inflorescences from section Phyllodineae (together with the bipinnate-leaved species of section Botrycephalae) to form a terminal monophyletic group, with the plurinerved species of sections Plurinerves and Juliflorae forming a series of basal paraphyletic groups. The close relationship between these racemose species of section Phyllodineae and species of section Botrycephalae has been noted by several authors (e.g. Tindale & Roux 1969 and 1974, Vassal 1972 and Pettigrew & Watson 1975, Chappill & Maslin 1995, Brain & Maslin 1996, Miller & Bayer 2001).
- Among the phyllode-bearing species there appears to be some sort of taxonomic discontinuity between at least certain ‘uninerved’ species of section Phyllodineae and the ‘plurinerved’ species of sections Plurinerves and Juliflorae. The classifications of Vassal (1972) and Pedley (1986) embody these affinities and there is some support for them in the results of recent studies involving morphology (Chappill & Maslin 1995) and DNA sequence data (Murphy et al. 2000, Miller & Bayer 2001, Miller et al. in press). Interestingly, however, the serological evidence of Brain & Maslin (1996) not only failed to support this ‘uninerved’ vs ‘plurinerved’ distinction but also suggested a positive relationship between certain non-racemose species of section Phyllodineae and species from sections Plurinerves and Juliflorae (see below). Clearly, much more study is needed to elucidate relationships between these higher-order groups within subgenus Phyllodineae. There are, for example, some “natural” groups which contain both uninerved and plurinerved species, e.g. the “A. wilhelmiana group” (Maslin 1990), A. murrayana (1-nerved phyllodes) - A. praelongata (plurinerved phyllodes) and the “A. paradoxa group” (where A. aspera, A. ausfeldii, A. dodonaeifolia, A. leprosa, A. paradoxa and A. rhetinocarpa have 1-nerved phyllodes whereas A. cognata, A. glandulicarpa, A. howittii, A. montanta, A. subporosa and A. verniciflua have plurinerved phyllodes). Also, diaphyllodinous species such as A. basedowii, A. diaphyllodinea, A. leptospermoides, etc. (see Maslin 1978 and Vassal & Maslin 1979) are anomalous in having phyllodes that are 1-nerved adaxially and 3-nerved abaxially.
- Among the plurinerved species there are examples of close relationships between species currently placed in section Juliflorae (inflorescences spicate) and section Plurinerves (inflorescences globular), for example, members of the A. stigmatophylla group (Tindale 1980); see also below under section Plurinerves. Additionally there are a number of species where the inflorescence shape is intermediate between a globular head and a cylindrical spike, e.g. species allied to A. filifolia (Chapman & Maslin 1999). This is not surprising because, as noted by Chappill & Maslin (1995), head shape is a plesiomorphic character in Acacia.
The serological investigation of subgenus Phyllodineae by Brain & Maslin (1996) revealed some higher-order patterns of relationships which were unexpected and which are at variance with established classifications. As already noted this study was not able to demonstrate a clear division between the uninerved species of section Phyllodineae and the plurinerved species of sections Juliflorae and Plurinerves. Furthermore, the serological evidence suggested that section Phyllodineae comprises two reasonably coherent subgroups, namely, species with elongated racemose inflorescences and those with non-racemose (or rudimentary racemose) inflorescences, and surprisingly there was no sign of a close relationship between the two subgroups. The non-racemose species of section Phyllodineae unexpectedly showed affinities with certain species currently included in sections Plurinerves and Juliflorae. If these relationships as indicated by the serological data are valid, then a major re-appraisal of the classification of subgenus Phyllodineae is needed.
Sections of Subgenus Phyllodineae
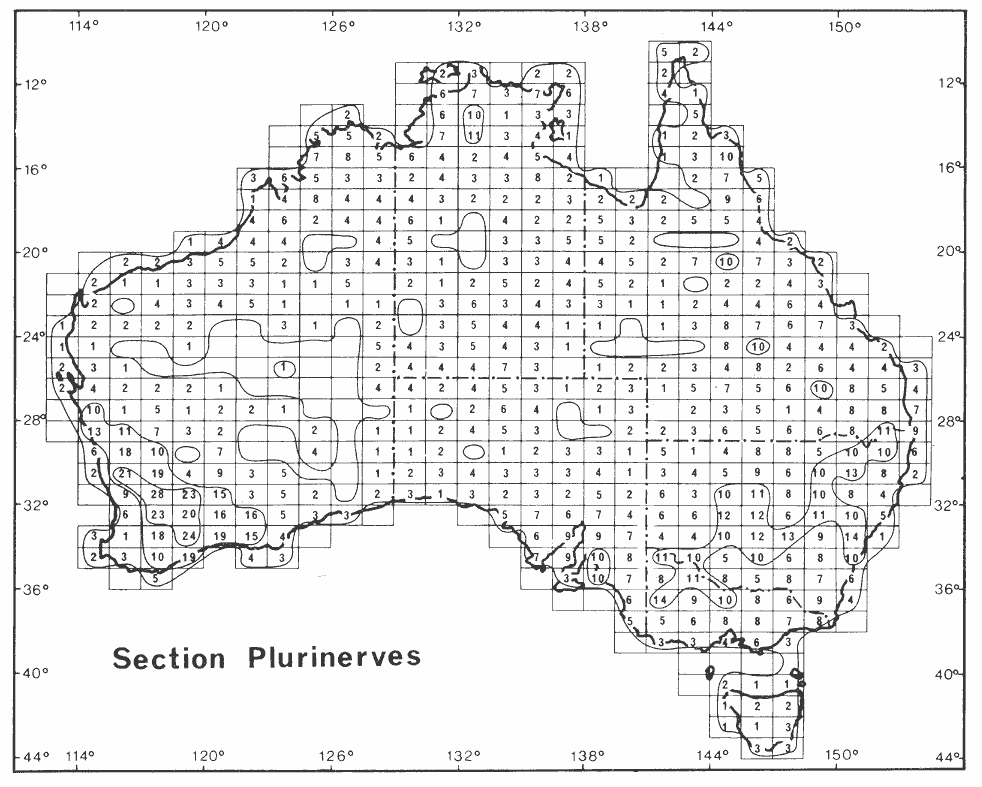
Section Plurinerves (216 described, accepted species). This large and diverse group contains species characterized by having plurinerved phyllodes and globular flower heads that are arranged in either simple or racemose inflorescences. Species of section Plurinerves are widespread in Australia with the main centres of richness located in the inland areas of the southwest and southeast of the continent (Hnatiuk & Maslin 1988, Maslin & Pedley 1988).
{kind=link}
A number of studies have suggested that section Plurinerves is related to section Juliflorae (see Tindale & Roux 1969 and 1974, Vassal 1972, Pettigrew & Watson 1975 and Tindale 1980) and indeed, in his most recent classification Pedley (1986) combined the two groups under Racosperma section Plurinerves. However, the cladistic analysis of Chappill & Maslin (1995) found that this group (i.e. the combined sections Plurinerves and Juliflorae) to be paraphyletic and that inflorescence shape, which has traditionally been used to separate the sections, was highly homoplastic. The results of recent molecular analyses by Miller & Bayer (2000, 2001), Murphy et al. (2000) and Miller et al. (in press) also show this group as paraphyletic. None of the above studies support section Plurinerves or section Juliflorae as monophyletic. The serological data of Brain & Maslin (1996) suggest a similar conclusion except that these authors did find some support for recognizing at least part of section Plurinerves as a ‘natural’ group. To resolve phylogenetic relationships involving these two sections more species need to be included in the on-going genetic investigations. Furthermore, elucidation of the complex variation patterns will be facilitated by developing a better understanding of phyllode nervature patterns.
In the absence of an effective infra-sectional classification of section Plurinerves a convenient (and somewhat natural) way of arranging the species is to group them according to their inflorescence structure (i.e. simple or racemose) and phyllode nervature (i.e. the number of longitudinal nerves, their spacing and whether or not intervening anastomosing secondary nerves are developed). Using these phyllode attributes, Pedley (1987a), Maslin & Pedley (1988) and Cowan & Maslin (1990a) recognized the following main nervation trends that apply to both section Plurinerves and section Juliflorae (however, not all species of the two sections can be conveniently accommodated within the groups that were defined):
- Phyllodes with numerous, very fine, close, longitudinal nerves without anastomoses between them (i.e. microneurous nervation).
- Phyllodes with few, widely-spaced longitudinal nerves (usually 3 nerves per face when phyllodes are flat, or 8-nerved in all when not flat); within this pattern two subgroups can be recognized.
- Anastomoses absent or few between the main longitudinal nerves.
- Anastomoses numerous between the main longitudinal (i.e. phyllodes reticulately veined).
The phyllode shape is moderately conservative, however they do vary from flat to terete/quadrangular. Notes on species groups within section Plurinerves are provided by Maslin & Stirton (1997).
View some examples of section Plurinerves taxa: A. dunnii, A. resinistipulea, A. resinosa.
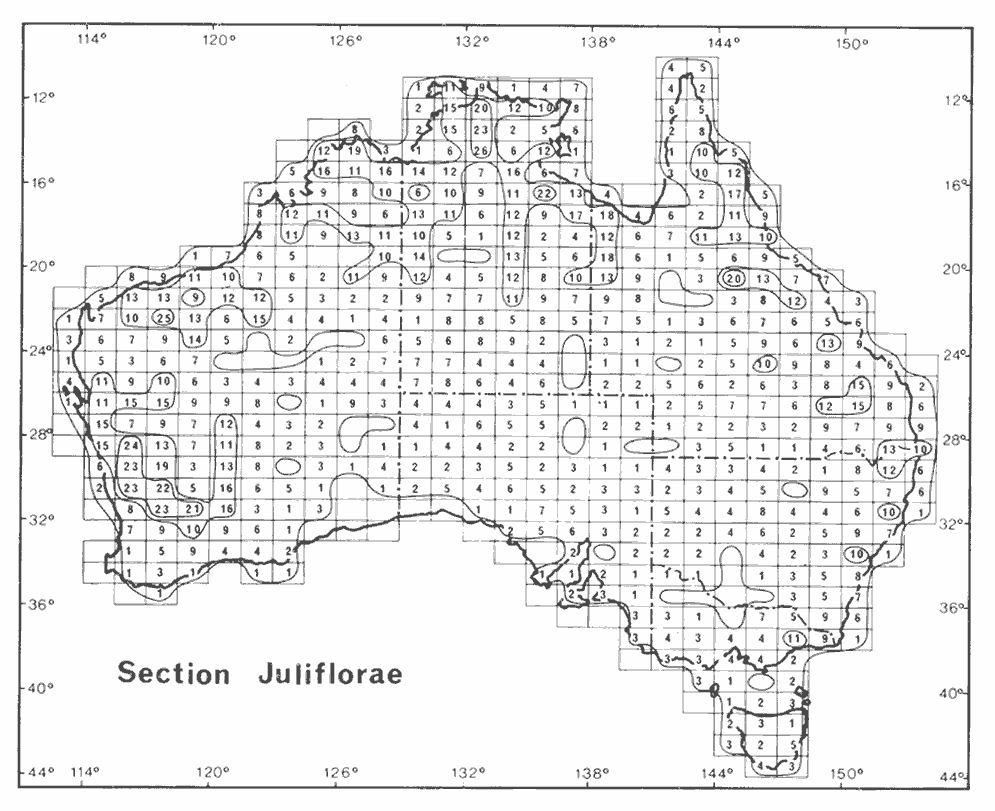
Section Juliflorae (255 described, accepted species). This large and diverse group contains species characterized by having plurinerved phyllodes and cylindrical flower heads that are arranged in either simple or racemose inflorescences. Species of section Juliflorae are widespread in Australia with the main centres of species richness occurring in the north, northwest and southwest of the continent and secondary centres of richness located along the Great Dividing Range in eastern Australia; although plants of this group often form a conspicuous element of the Arid Zone flora, species numbers in these areas are generally not great (Hnatiuk & Maslin 1988, Maslin & Pedley 1988). This geographic pattern is similar to that of section Plurinerves, however, there are differences between the two group. The most important of these being that section Plurinerves has fewer species north of the Tropic of Capricorn and more species in inland areas of south-eastern Australia (see Hnatiuk & Maslin 1988); a few species of both sections occur outside Australia (Pedley 1975).
{kind=link}
As already noted above section Plurinerves is more closely related to section Juliflorae than either is to section Phyllodineae. The precise nature of this relationship is as yet unresolved, however, it is not likely that either section, or the two sections combined, are monophyletic. Traditionally section Juliflorae and Plurinerves are separated on the basis of the shape of their inflorescences, cylindrical in the former and globular the latter section, however, Chappill & Maslin (1995) found this to be a highly homoplastic character.
Currently there is no effective classification of species included in section Juliflorae. However, as with section Plurinerves phyllode nervature is helpful arranging species into groups. Inflorescence structure is less important for grouping the species than it is in section Plurinerves, however, details of the flower (especially the degree of calyx division) and pod morphology are useful attributes. The phyllode shape is moderately conservative, however they do vary from flat to terete/quadrangular. Notes on species groups within section Juliflorae are provided by Maslin & Stirton (1997).
View some examples of section Juliflorae taxa: A. cowleana, A. elachantha, A. neurophylla subsp. erugata, A. stereophylla subsp. stereophylla, A. hilliana.
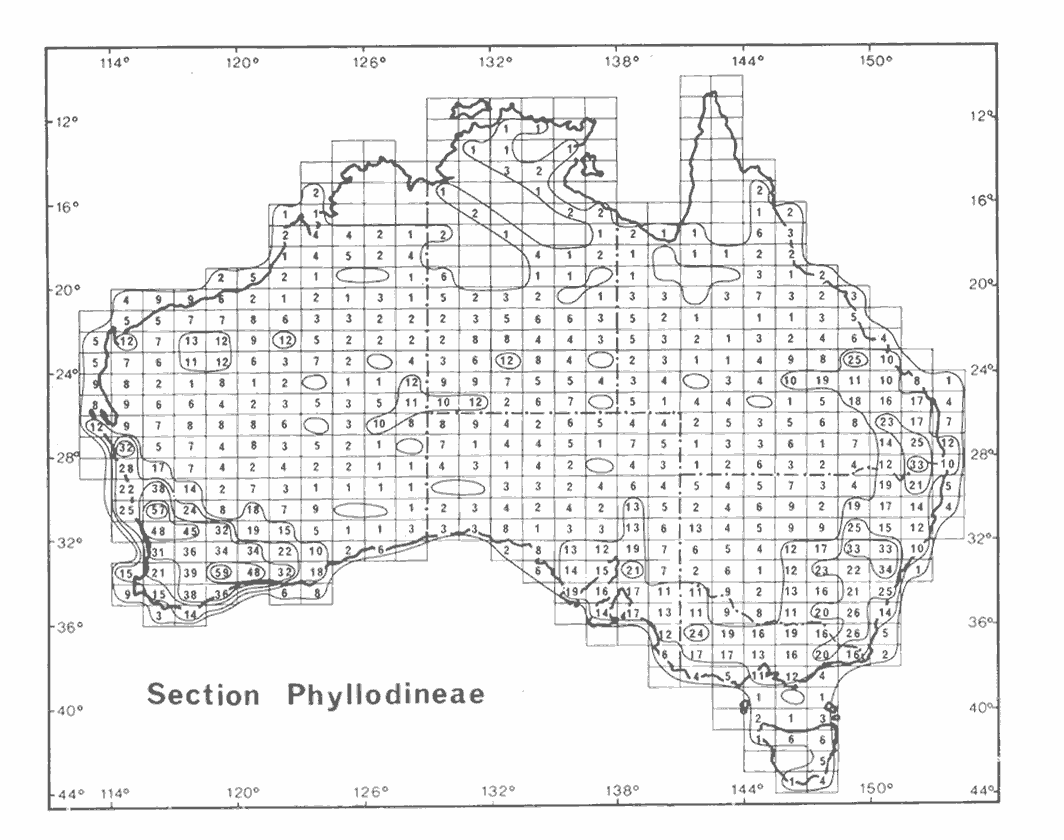
Section Phyllodineae(397 described, accepted species). This is the largest group within subgenus Phyllodineae. The species are recognized by their ‘1-nerved’ phyllodes and their normally globular flower heads arranged in either simple or racemose inflorescences. The section is widespread in Australia with the most species-rich areas occurring in southwest Western Australia and south of the Tropic of Capricorn in the temperate and adjacent semiarid areas of eastern and south-eastern Australia; the section is poorly represented in the Arid Zone and virtually absent from the tropical north of the continent (Hnatiuk & Maslin 1988, Maslin & Pedley 1988). There are no indigenous species of section Phyllodineae that occur outside Australia.
{kind=link}
In the absence of an effective classification of species within section Phyllodineae a convenient (albeit somewhat oversimplified) way to arrange them is to divide them into two groups based on inflorescence structure, namely, racemose vs non-racemose. Serological evidence presented by Brain & Maslin (1996) suggest that fundamental differences exist not only between these two groups, but also between species-complexes within each group. These results are supported by recent molecular evidence by Miller & Bayer (2000, 2001), Murphy et al. (2000) and Miller et al. (in press). These studies tend to negate the assertion of Chappill & Maslin (1995) that section Phyllodineae is monophyletic. Furthermore, the results of the Brain & Maslin (1996) study (surprisingly) suggested that that some of the non-racemose species of section Phyllodineae have their greatest affinities with plurinerved species of sections Juliflorae and Plurinerves.
Although Bentham (1864) recognized six subseries within his series Uninerves (= section Phyllodineae) these are largely artifical groups and are generally not adopted by modern workers. The phyllodes on species of section Phyllodineae are more variable than those of sections Juliflorae and Plurinerves; they vary from flat to terete/quadrangular and sometimes display rather bizarre shapes (especially in southwest Western Australia). Notes on species groups within section Phyllodineae are provided by Maslin & Stirton (1997).
View some examples of section Phyllodineae taxa: A. delphina, A. inaequilatera, A. suaveolens.
Section Botrycephalae (40 described, accepted species). This group comprises mostly arborescent species that are characterized by having bipinnate adult foliage and normally elongated racemose inflorescences; it predominates in temperate areas of eastern and southeastern Australia (Hnatiuk & Maslin 1988, Maslin & Pedley 1988). As discussed above there is a considerable body of evidence that shows a close relationship exists between section Botrycephalae and certain phyllodinous species (possessing racemose inflorescences) from section Phyllodineae.
{kind=link}
Relationships between species currently ascribed to section Botrycephalae have not yet been fully elucidated, however, based on chemical data a number of subgroups have been recognized. Studies by Tindale and Roux (1969) of flavonoid and condensed-tannin contents of the heartwood and bark from l8 species recognized the following species-groups: (1) A. constablei, A. decurrens, A. irrorata subsp. velutinella, A. mearnsii, A. parramattensis and A. trachyphloia; (2) A. filicifolia, A. irrorata and A. silvestris; (3) A. botrycephala, A. cardiophylla, A. chrysotricha, A. leucoclada subsp. argentifolia and A. oshanesii; (4) A. baileyana and A. dealbata. Also, from detailed analyses of gum from 10 species Anderson (1978) and Anderson et al. (1971 and 1984) proposed that section Botrycephalae be divided into two distinct groups. Group A included six taxa (A. deanei and A. deanei subsp. paucijuga, A. irrorata subsp. irrorata, A. parramattensis, A. parvipinnula and A. trachyphloia) which differed greatly in their gum composition from the phyllodinous species of Bentham's (1864) “Division” Phyllodineae; Group B also contained six taxa (A. dealbata subsp. dealbata and subsp. subalpina, A. filicifolia, A. leucoclada, A. silvestris and A. terminalis) but these showed a strong chemical resemblance to the phyllodinous taxa examined. Pedley (1986) suggested that perhaps A. leptoclada and A. mitchellii might constitute a third group. Acacia mitchellii, which differs from all other species of Botrycephalae in having non-racemose inflorescences, was excluded from section Pulchellae by Guinet et al. ( 1980).
View some examples of section Botrycephalae taxa: A. parramattensis, A. mearnsii.
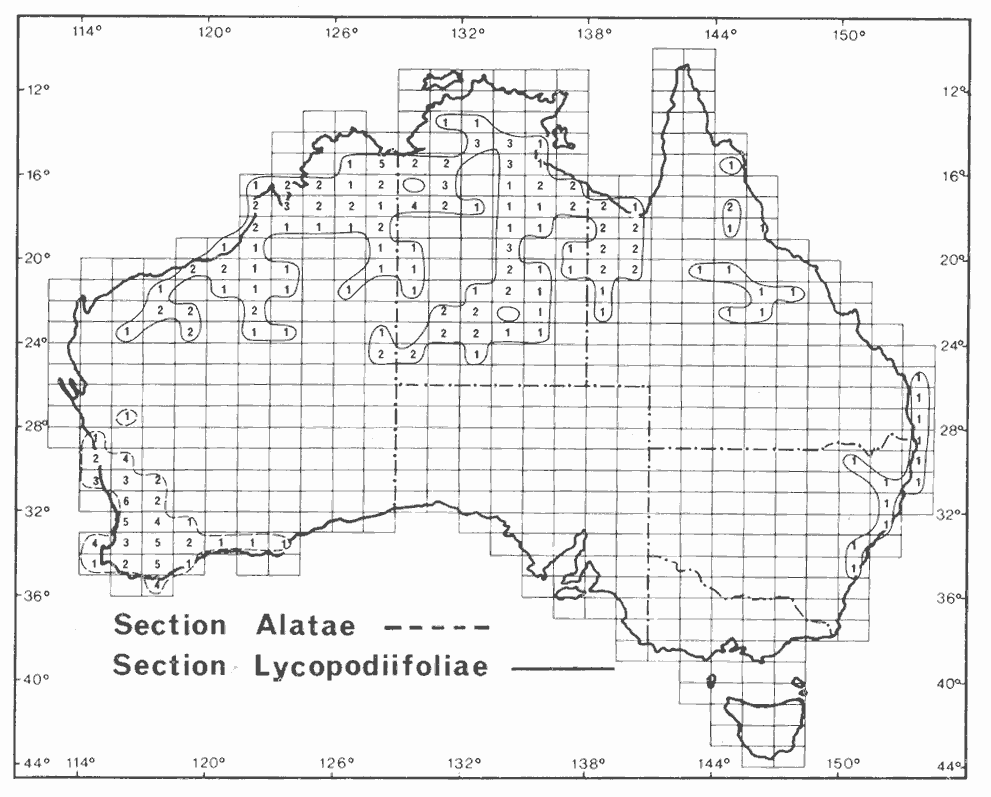
Section Alatae (c. 23 described, accepted species). Although Pedley (1978) recognized this section, it is a polyphyletic assemblage (Chappill & Maslin 1995, Murphy et al. 2000) and is unlikely to be recognized in future classifications. The decurrent phyllodes which define this group were interpreted by Chappill & Maslin (1995) as a highly homoplastic feature.
{kind=link}
View some examples of section Alatae taxa: A. alata var. alata, A. continua.
Section Lycopodiifoliae (18 described, accepted species). This distinctive group is characterized by having whorled phyllodes and is found in the Arid Zone and in northern areas of the continent (see Hnatiuk & Maslin 1988 and Maslin & Pedley 1988).
Section Lycopodiifoliae probably constitutes a monophyletic group (Murphy et al. 2000) but its relationship to the rest of subgenus Phyllodineae is not clear. Pedley (1987a) and Cowan & Maslin (1990) suggested affinities A. deltoidea and its allies (section Plurinerves) and this relationship warrants further investigation. The monophyletic group recognized by Murphy et al. (2000) containing species of section Lycopodiifoliae and section Pulchelloidea (Vassal 1972) is difficult to reconcile based on morphological criteria. The character of verticillate phyllodes which characterizes this section has arisen independently in a number of unrelated groups of Acacia, e.g. A. cedroides and A. verticillata.
View some examples of section Lycopodiifoliae taxa: A. adoxa var. adoxa, A. spondylophylla.
Section Pulchellae (26 described, accepted species). This group of bipinnate-leaved species is endemic in south-west Western Australia (see Hnatiuk & Maslin 1988 and Maslin & Pedley 1988). This section is reviewed by Maslin (1975, 1979) and as discussed by Guinet et al. (1980) it comprises a “core” of interrelated species centered around A. pulchella, A. browniana and A. drummondii. Some of these species, together with certain phyllodinous taxa, were included by Vassal (1972) in his section Pulchelloidea. Although section Pulchelloidea was not supported by Pettigrew & Watson (1975) there is recent molecular evidence by Murphy et al. (2000) which suggests that it warrants further investigation. As already noted, however, the inclusion by Murphy et al. (2000) of species referable to section Pulchelloidea and section Lycopodiifoliae in the one monophyletic group is difficult to reconcile by morphological criteria.
View some examples of section Pulchellae taxa: A. drummondii subsp. drummondii, A. pulchella var. reflexa.
References
Anderson, D.M.W. (1978). Chemotaxonomic aspects of the chemistry of Acacia gum exudates. Kew Bulletin 32: 529–536.
Anderson, D.M.W., Bell, P.C. and McNab, C.G.A. (1971). An analytical study of some Acacia gum exudates of the series Botryocephalae. Carbohydrate Research 20: 269–274.
Anderson, D.M.W., Farquhar, J.G.K. and McNab, C.G.A. (1984). The gum exudates from some Acacia subspecies of the series Botryocephalae. Phytochemistry 23: 579–580.
Bentham, G. (1842) Notes on Mimoseae, with a Synopsis of Species. London Journal of Botany 1: 318–528.
Bentham, G. (1864) Flora Australiensis. vol. 2. (Lovell Reeve, London.)
Bentham, G. (1875) Revision of the Suborder Mimoseae. Trans. Linn. Soc. London 30: 335–664.
Brain, P. and Maslin, B.R. (1996). A serological investigation of the classification of Acacia subgenus Phyllodineae (Leguminosae: Mimosoideae). Biochemical Systematics and Ecology 24: 379–392.
Chapman, A.R. and Maslin, B.R. (1999). Acacia miscellany 20. Descriptions of three new Western Australian species of Acacia section Juliflorae (Leguminosae: Mimosoideae). Nuytsia 12(3): 487–491.
Chappill J.A. and Maslin, B.R. (1995). A phylogenetic assessment of tribe Acacieae. In Advances in Legume Systematics 7: Phylogeny. (Eds M.D. Crisp and J.J. Doyle ) pp. 77–99. (Royal Botanic Gardens, Kew: London)
Cowan, R.A. (1998). Mimosaceae (excl. Acacia), Caesalpiniaceae. Flora of Australia vol. 12 (ABRS/CSIRO Publishing).
Cowan, R.S. and Maslin, B.R. (1990). Acacia Miscellany 2. Species related to A. deltoidea (Leguminosae: Mimosoideae: section Plurinerves). Nuytsia 7: 201–208.
Cowan, R.S. and Maslin, B.R. (1990a). Acacia Miscellany 1. Some oligoneurous species of Acacia (Leguminosae: Mimosoideae: Section Plurinerves) from Western Australia. Nuytsia 7: 183–199.
Elias, T.S. (1981). Mimosoideae. In Advances in Legume Systematics. Part 1. (Eds. R.M. Polhill and P.H. Raven) pp. 143–151. (Royal Botanic Gardens, Kew: London.)
Grimes, J.W. (1999). Inflorescence morphology, heterochrony, and phylogeny in the Mimosoid tribes Ingeae and Acacieae (Leguminosae: Mimosoideae). Botanical Review 65: 317–347.
Guinet, P. and Vassal, J. (1978). Hypotheses on the differentiation of the major groups in the genus Acacia (Leguminosae). Kew Bulletin 32: 509–527.
Guinet, P., Vassal, J., Evans, C.S. and Maslin, B.R. (1980). Acacia (Mimosoideae): composition and affinities of the series Pulchellae Bentham. Botanical Journal of the Linnean Society 80: 53–68.
Hnatiuk, R.J. and Maslin, B.R. (1988). Phytogeography of Acacia in Australia in relation to climate and species-richness. Australian Journal of Botany 36: 361–383.
Jawad, J.T., Seigler, D.S. and Ebinger, J.E. (2000). A systematic treatment of Acacia coulteri (Fabaceae, Mimosoideae) and similar species in the New World. Annals of the Missouri Botanical Garden 87: 528–548.
Luckow, L., Miller, J.T., Murphy, D.J. and Livshultz, T. (in press). A phylogenetic analysis of the Mimosoideae (Leguminosae) based on chloroplast DNA sequence data. In Advances in Legume Systematics. (Eds B Klitgaard and A Bruneau). (Royal Botanic Gardens: Kew, London)
Maslin, B.R. (1975). Studies in the genus Acacia (Mimosaceae) 4. A revision of the series Pulchellae. Nuytsia 1: 388–492.
Maslin, B.R. (1978). Studies in the genus Acacia (Mimosaceae) - 7. The taxonomy of some diaphyllodinous species. Nuytsia 2: 200–219.
Maslin, B.R. (1979). Studies in the genus Acacia (Mimosoideae) – 9. Additional notes on the Series Pulchellae Benth. Nuytsia 2: 354–367.
Maslin B.R. (1987). Report of discussions at I.G.S.M. meeting no. 6 on the proposal to split Acacia. Bulletin of the International Group for the Study of Mimosoideae 15: 108–118.
Maslin, B.R. (1988). Should Acacia be divided? Bulletin of the International Group for the Study of Mimosoideae 16: 54–76. (This paper was reproduced under a different title in 1989: see next entry.)
Maslin B.R. (1989). Wattle become of Acacia? Australian Systematic Botany Society Newsletter 58: 1–13.
Maslin, B.R. (1990). Acacia Miscellany 4. Three new Western Australian species with affinities to A. wilhelmiana (Leguminosae: Mimosoideae: section Plurinerves) from Western Australia. Nuytsia 7(2): 221–228
Maslin, B.R. (2001). Introductory essay. In A.E. Orchard & A.J.G. Wilson (eds) Flora of Australia Volume 11A, Mimosaceae, Acacia part 1, pp. 3–13. (ABRS/CSIRO Publishing.)
Maslin, B.R., Byrne, M., Coates, D., Broadhurst, L., Coleman, H. and Macdonald, B. (1999). The Acacia acuminata (Jam) group: an analysis of variation to aid Sandalwood (Santalum spicatum) plantation research. (Unpublished report to the Sandalwood Business Unit, Department of Conservation and Land Management: Perth.)
Maslin, B.R., Miller, J.T. and Seigler, D.S. (2003). Overview of the generic status of Acacia (Leguminosae: Mimosoideae). Australian Systematic Botany 16(1): 1–18. An abstract of this paper (together with a provision to purchase the whole article) is available from CSIRO Publishing.
Maslin, B.R. and Pedley, L. (1988). Patterns of distribution of Acacia in Australia. Australian Journal of Botany 36: 385–393.
Maslin, B.R. and Stirton, C.H. (1997). Generic and infrageneric classification in Acacia (Leguminosae: Mimosoideae): a list of critical species on which to build a comparative data set. Bulletin of the International Group for the Study of Mimosoideae 20: 22–44.
Miller, J.T., Andrew, R. and Bayer, R.J. (in press). Molecular Phylogenetics of the Australian Acacias of subg. Phyllodineae (Fabaceae: Mimosoideae) based on the trnK intron. (To be published in Australian Journal of Botany.)
Miller J.T. and Bayer R.J. (2000). Molecular phylogenetics of Acacia (Fabaceae: Mimosoideae) based on chloroplast TRNK/MATK and nuclear histone H3-D sequences. In Advances in Legume Systematics 9. (Eds PS Herendeen and A Bruneau ) pp. 180–200 (Royal Botanic Gardens, Kew: London)
Miller J.T. and Bayer R.J. (2001). Molecular phylogenetics of Acacia (Fabaceae: Mimosoideae) based on the chloroplast MATK coding sequence and flanking TRNK intron spacer region. American Journal of Botany 88: 697–705.
Miller J.T. and Bayer R.J. (2003). Molecular phylogenetics of Acacia subgenera Acacia and Aculeiferum (Fabaceae: Mimosoideae) based on the chloroplast matK coding sequence and flanking trnK intron spacer regions. Australian Systematic Botany 16: 27–33.
Miller, P. (1754). The Gardeners' Dictionary. Abr. 4th ed. London.
Murphy, D.J., Udovicic, F, and Ladiges, P.Y. (2000). Phylogenetic analysis of Australian Acacia (Leguminosae: Mimosoideae) by using sequence variations of an intron and two intergenic spacers of chloroplast DNA. Australian Systematic Botany 13: 745–754.
Orchard, A.E. and Maslin, B.R. (2003). Proposal to conserve the name Acacia (Leguminosae: Mimosoideae) with a conserved type. Taxon 52(2): 362–363. A web download of this paper is available.
Pedley, L. (1975). Revision of the extra-Australian species of Acacia subg. Heterophyllum. Contributions from the Queensland Herbarium 18: 1–24.
Pedley, L. (1978). A revision of Acacia Mill. in Queensland. Austrobaileya 1: 75–234.
Pedley, L. (1986). Derivation and dispersal of Acacia (Leguminosae), with particular reference to Australia, and the recognition of Senegalia and Racosperma. Botanical Journal of the Linnean Society Soc. 92: 219–254.
Pedley, L. (1987). In defence of Racosperma. Bulletin of the International Group for the Study of Mimosoideae 15: 123–129.
Pedley, L. (1987a). Racosperma deltoideum (Cunn. ex G. Don) Pedley (Leguminosae: Mimosoideae) and related species on northern Australia. Austrobaileya 2(4): 314–320.
Pedley, L. (1989). Racosperma again. Australian Systematic Botany Newsletter 59: 1–2.
Pettigrew, C.J. and Watson, L. (1975). On the classification of Australian Acacias. Australian Journal of Botany 23: 833–347.
Polhill, R.M. (1994). Classification of the Leguminosae. In: F.A. Bisby, J. Buckingham, and J.B. Harborne (eds) Phytochemical Dictionary of the Leguminosae, vol 1. Plants and their constituents, pp. 35-36 (Chapman & Hall: London.)
Robinson, J. and Harris, S.A. (2000). A plastid DNA phylogeny of the genus Acacia Miller (Acacieae, Leguminosae). Botanical Journal of the Linnean Society 132: 195–222.
Tindale, M.D. (1980). Notes on Australian taxa of Acacia No. 7. Telopea 2: 113–125.
Tindale, M.D. and Roux, D.G. (1969). A phytochemical survey of the Australian species of Acacia. Phytochemistry 8: 1713–1727.
Tindale, M.D. and Roux, D.G. (1974). An extended phytochemical survey of Australian species of Acacia: chemotaxonomic and phylogenetic aspects. Phytochemistry 13: 829–839.
Vassal, J. (1972). Apport des recherches ontogéniques et seminologiques à l'étude morphologique, taxonomique et phylogénique du genre Acacia. Bulletin de la Societe d’Histoire Naturelle de Toulouse 108: 105–247.
Vassal, J. and Maslin, B.R. (1979). Phyllodes et phyllodisation das le genre Acacia. Bulletin de la Societe d’Histoire Naturelle de Toulouse 115: 393–401.